白斑综合征病毒WSSV(white spot syndrome virus)是全世界范围内甲壳类动物的重要病原,至今未获得对其有效的防治措施,因此开发抗WSSV药物对于甲壳类动物养殖产业的健康可持续发展具有重要意义。WSSV基因组能够编码胸苷酸合成通路中的关键酶dUTP焦磷酸酶(dUTP pyrophosphatase,dUTPase),该酶存在于绝大多数生物中,催化dUTP水解产生dUMP和焦磷酸PPi,对于DNA的合成与稳定起着不可或缺的作用,是抗病原体药物开发的重要靶标。开发以dUTPase为靶标的抗WSSV药物,需要深入了解WSSV和宿主dUTPase的结构特点。
本文通过解析WSSV和凡纳滨对虾Litopenaeus vannamei(WSSV侵染的主要养殖对象)dUTPase保守结构域的晶体结构,结合氨基酸定点突变和酶动力学分析等方法,深入探究两者的结构与功能特点,阐明其结构差异,不但从分子水平丰富了对dUTPase家族分类和催化反应机制的认识和理解,并且为抗WSSV药物的开发提供了准确的蛋白结构信息。
氨基酸序列比对显示,WSSV基因组中wsv112基因编码的蛋白序列N端171个氨基酸(wDUT)构成的dUTPase属于三聚体家族,包含酶催化活性必需的5个基序(I-V)。wDUT在基序V前具有特异性插入序列pre-V,该序列由20个氨基酸(Tyr134-Asn153)组成。wDUT晶体结构分析发现,pre-V形成独特的β发夹结构,引导其后C端序列产生了新颖的走向,导致基序V覆盖到自身亚基参与形成的底物结合口袋上,从而使wDUT活性位点仅由两个亚基组成,这与三聚体dUTPase活性位点由三个亚基组成的传统认识截然不同。该结果表明wDUT是同源三聚体dUTPase家族中新的亚型,扩充了对dUTPase家族分类的认识。
通过结构比对和氨基酸定点突变体的酶动力学分析发现,wDUT活性位点的氨基酸组成和催化反应机制与传统型三聚体dUTPase基本一致。然而,pre-V截短后,酶催化效率及蛋白热稳定性会显著下降,这表明pre-V截短体中由三个亚基形成的活性位点并不具有高水平酶催化效率,暗示wDUT发生了适应于新型活性位点组织方式的氨基酸替代。进一步利用wDUT和大肠杆菌dUTPase(传统型三聚体代表)突变体进行酶动力学分析发现,传统型三聚体dUTPase基序IV中保守Arg/Lys的功能被wDUT基序I前特异的Arg24替代,并且该wDUT活性位点周围氨基酸的重排与其新颖的C端走向相适应,共同维持了wDUT高水平的酶催化效率,该结果表明wDUT发生了序列插入与氨基酸突变的协同进化。
不同于WSSV的wDUT蛋白,宿主凡纳滨对虾dUTPase序列C端的146个氨基酸(LvDUT)为传统的同源三聚体dUTPase保守结构域。通过LvDUT多种反应状态晶体结构的比较,我们首次发现底物dUTP的结合能够引起dUTPase蛋白主体部分产生两种新型的构象变化:一是活性位点区域Ser132-His138由无规则卷曲变为α螺旋,二是三聚体中央通道一端发生“开合”。这两种构象变化共同促进了dUTP水解反应的发生。
wDUT和LvDUT结构比对表明,两者在传统的dUTPase抑制剂设计位点,即底物结合口袋和三聚体中央通道上差异较小,不利于针对wDUT抑制剂的开发和使用。但是,在wDUT独特的C端走向上,Lys150-Asn155与蛋白主体形成的空间中存在合适的小分子化合物结合位点,可以作为wDUT潜在的特异性抑制剂结合靶位,这为抗WSSV特异性药物的开发奠定了分子结构基础。 | 

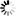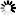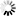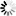

修改评论